GmDREB6 Geninin Soya Fasülyesine Ekspresyonu, Genetik Olarak Modifiye Edilmiş Soya Bitkilerinde Prolin Birikimini ve Tuz Toleransını Arttırıyor

Soya fasulyesi bitkileri abiyotik stresin etkilerine duyarlı ve daha az kurak-tuz toleransı olan mahsul grubuna aittir. Su sıkıntısıyla başa çıkmayı hedefleyen mekanizmalarda yer alan genlerin belirlenmesi, soya fasulyesinin kuraklık ve tuz toleransını arttırmak için vazgeçilmez bir görevdir. Daha fazla toleranslı mahsül elde etmek için bu konudaki yaklaşımlardan biri genetik modifikasyondur. AP2 familyasına ait dehidrasyona duyarlı elemanı bağlayıcı proteinler (DREB’ler), kuraklığa ve tuza toleranslı tepkilere aracılık eden hedef genlerin ekspresyonunu aktive etmek için promotörün cis sekanslarına bağlanan trans-aktif transkripsiyon faktörleridir. Bu çalışmada; GmDREB6 transgeni, Agrobacterium aracılığıyla transformasyon ile DT84 çeşidi soya fasulyesi bitkilerine eklenmiştir. Genetik olarak modifiye edilmiş (GM) soya fasulyesi bitkilerinde transkripsiyonel GmP5CS seviyesinin ve prolin birikiminin arttırılmasında GmDREB6 aşırı ekspresyonunun etkinliği de analiz edildi. Sonuçlar, kanamisin ile seçildikten sonra dönüştürülmüş eksplantlardan 10 GM soya fasulyesi bitkisinin (T0 jenerasyonu) başarıyla üretildiğini gösterdi. Bu bitkiler arasında, GmDREB6 transgeninin mevcudiyeti, dokuz bitkide Polimeraz Zincir Reaksiyonu (PCR) ile teyit edildi ve sekiz bitki, Southern blot’ta pozitif sonuçlar gösterdi. T1 neslinde; T1-2, T1-4, T1-7 ve T1-10 olarak etiketlenmiş dört GM hattı, rekombinant GmDREB6 proteinini temsil ediyor. T2 generasyonunda, GmP5CS geninin transkripsiyon seviyeleri, GM hatlarında, normal koşullar altında ve ayrıca tuz stresi ve kuraklık koşulları altında, 1.36 ila 2.01 kat ve 1.58 ila 3.16 kat arasında değişmekte ve transgenik olmayan bitkilerdir. Prolin içeriği; dört GM soya fasulyesi hattında (T2-2, T2-4, T2-7 ve T2-10), 0.82 μmol / g ile 4.03 μmol / g arasında değişen transgenik olmayan bitkilerde daha yüksek. Prolin içeriği GM T2-7 çizgisinde (7.77 µmol / g) en yüksek. GM soya fasulyesi hatlarında, bitkiler normal şartlara kıyasla yedi gün boyunca tuz stresine maruz bırakıldıktan sonra T2-2, T2-4, T2-7 ve T2-10 ; prolin içeriğinin arttığını ve % 247.83 ila 300 arasında değiştiğini, GM olmayan bitkilerinki değerlerinin ise % 238.22 olduğu gözlendi. Bu elde edilen sonuçlar, GmDREB6’nın tuz gibi bir abiyotik strese karşı toleransı artırmak için genetik mühendisliği için potansiyel bir aday olabileceğini düşündürmektedir.
Soya fasulyesi, sayısız kullanıma sahip önemli bir üründür; insan gıdası ve hayvan yemi endüstrisi için bir protein ve bitkisel yağ kaynağıdır. Soya ekimi toprağın ekonomik değerini, besleyici değerini ve verimliliğini artırır. Bununla birlikte, diğer mahsuller gibi, soya fasulyesi bitkileri de çevresel koşullara maruz kalmakta ve kuraklık, aşırı tuzluluk dahil olmak üzere abiyotik streslerin etkileriyle karşı karşıyadır. Kuraklık stresi ve uzun süreli kuraklığın soya fasulyesi bitkilerinin büyümesi ve gelişmesi üzerinde çok büyük etkileri vardır; örneğin kuraklık tahıl oluşumu aşamasında ortaya çıkarsa, verimi ve tohum kalitesini yaklaşık % 40 oranında azalır. Soya mahsullerinin kuraklığa ve tuz stresine karşı dayanıklılığının iklim değişikliğine adaptasyon olduğu düşünülmektedir. Kuraklık ve tuz streslerinin soya fasulyesi bitkileri üzerindeki olumsuz etkilerini en aza indirmek için; bu stres koşullarına daha toleranslı olan genetik olarak değiştirilmiş (GM) çeşitlerin yanı sıra, sulama sistemlerinin iyileştirilmesi ve toprakta nemin tutulmasının iyileştirilmesi gibi konularda da alternatif stratejiler kullanılabilir. . Ayrıca, kuraklığa ve tuz toleranslı soya türlerinin çok aranmasının bir nedeni; yüksek bölgelerin kuraklık ve tuzluluk oranın iklim değişikliği sebebiyle gelecekte artacağı tahmin edilmektedir. Hem kuraklık hem de tuz stresi, ozmotik strese ve sitoplazmadan suyun çıkarılmasına ve sonuç olarak sitosolik ve vakuolar hacimlerin azalmasına neden olan hücresel dehidrasyona yol açar.
Genetik mühendisliği, kuraklığa ve tuza toleranslı ürünlerin geliştirilmesi için güçlü bir araçtır, çünkü kuraklık ve tuz streslerine yanıt olarak gen ifadesinin değiştirilmesi ile istenen özelliklere sahip ürünler elde edilir. Bu durumda, GM ve tuza dayanıklı soya fasulyesi bitkilerinin gelişimi için su sıkıntısı ve yüksek tuzluluk ile başa çıkmayı hedefleyen mekanizmalarda yer alan genlerin belirlenmesi esastır. Kuraklığa yanıt olarak gen ekspresyonu, transkripsiyon mekanizmaları ve sinyal iletiminin anlaşılmasındaki gelişmeler daha önce incelenmiştir. Kuraklık tepkilerinde yer alan genler arasında, transkripsiyon faktörlerini (TF’ler) kodlayan genler, bitkilerin genetik mühendisliği için arzu edilen adaylarıdır, çünkü TF’ler hedef genlerin promotör bölgesindeki spesifik DNA dizilerini tanır ve ekspresyonu düzenler. Bu şekilde hareket eden TF’ler, bazı kuraklık ve tuz stresine duyarlı genlerin ekspresyonunu düzenler. Bazı çalışmalar, TF’lerin GM soya fasulyesi bitkilerinin kuraklık ve tuz toleransını iyileştirmedeki rolünü değerlendirmeyi sağlamıştır. DREB’ler (dehidrasyona duyarlı eleman bağlayıcı proteinler), su eksikliği ile transkripsiyonel olarak bitkiyi regüle eden TF’lerdir. AP2 familyasına ait soya fasulyesindeki DREB’ler, hedef genlerin promotör bölgesindeki cis sekanslarına bağlanan, böylece dış ortamdan abiyotik stres sinyallerine yanıt olarak ekspresyonlarını aktive eden trans-aktif TF’lerdir. AP2 alanı yaklaşık 58 veya 59 amino asit içerir ve dehidrasyon tepki faktörüne (DRE) veya GCC kutusuna bağlı 8 amino asitten oluşur.
Bazı çalışmalar, DREB genleri içeren bitkilerin serada veya tarla koşullarında abiyotik streslere karşı toleransı arttırdığı gözlemlenmiştir. Bu çalışmalar ayrıca, daha iyi kuraklık toleransı tepkileri nedeniyle GM bitkilerinin veriminin kuraklık koşullarında sabit kaldığını ortaya konmuştur. AtDREB1A, AtDREB2CA ve AtAREB GM soya fasulyesi hatları, sulanan ve sulanmayan koşullar altındaki bir alanda analiz edildi. Çalışma; tarla deneyinde kuraklık koşulları sırasında GM bitkilerinin kuraklığa daha dayanıklı olduğunu doğruladı. Bu çalışmalar DREB1 ve DREB2 genlerini ve verim bileşenleri, agronomik özellikler ve fizyolojik parametreler üzerinde yoğunlaşmıştır. GmDREB6 geni için, kuraklık ve tuz toleransının iyileştirilmesi ve GmDREB6’yı aşırı eksprese eden GM soya bitkilerinde analiz edilen prolin içeriği ile ilgili hiçbir çalışma yapılmamıştır. Prolin, kuraklık koşullarında bitkilerde artan yaygın bir ozmolittir ve birikmesi ozmotik basıncı arttırır, böylece bitkilerin tuz toleransıda artar. Zhang ve arkadaşları; OsDREB2A’nın ekspresyon sonuçlarını inceledi ve ayrıca bu genin ekspresyonunun soya fasulyesinde DREB6 ve P5CS gen ekspresyonunu arttırdığını keşfetti. Bununla birlikte, soya fasulyesi DREB6’nın aşırı ekspresyonunun P5CS ekspresyonunu nasıl arttırdığına dair hiçbir veri veya prolin içeriği bu çalışmada test edilmiştir. Bu bağlamda çalışma; GmDREB6 transgeninin ekspresyonunu ve GmDREB6 aşırı ekspresyonunun, yüksek tuzluluk koşulları altında GM soya fasulyesi bitkilerinde GmP5CS geninin transkripsiyon seviyelerinin ve prolin birikiminin arttırılmasındaki etkinliğini analiz etmeyi amaçlamıştır.
35S-GmDREB6-c-myc ; yapı-pozitif GM bitkilerinin tanımlanması DT84 soya fasulyesi çeşidinden dört yüz elli kotiledon düğümü, Agrobacterium tumefaciens aracılı transformasyon kullanılarak 35S-GmDREB6-c-myc genetik yapısı ile transforme edildi . Bir kontrol düzeneği olarak, transgenik olmayan soya fasulyesi kotiledonları antibiyotik içermeyen bir ortamda yeniden üretildi ve 8 transgenik olmayan soya fasulyesi bitkisi seçildi ve seradaki saksılara aktarıldı.

Fotoğraf : Soya fasulyesi bitkilerinin transformasyonu ve in vitro rejenerasyonu. (A): Kotiledonlar, pBI121 :: GmDREB6 vektörünü 30 dakika taşıyan A. tumefaciens ile inkübe edilerek enfekte edildi; (B): CCM ile karanlıkta 5 gün boyunca birlikte kültivasyon; (C): Kotiledonlar, 2 hafta boyunca 2 mg / L BAP ve 50 mg / L kanamisin ile muamele edilmiş SIM çoklu sürgün yenileme ortamında kültürlendi; (D): Kotiledonlar çıkarıldı ve 2 hafta boyunca 0.5 mg / L GA3, 0.1 mg / L IAA ve 50 mg / L kanamisin ile desteklenen SEM üzerinde kültürlendi; (E): 20 gün boyunca 0.1 mg / L IBA ile takviye edilmiş RM’deki yenilenmiş sürgünlerde kök büyümesinin başlatılması; (F): Köklü bitkiler, 1: 1 oranında pirinç kabuğu kömürü ve kum karışımı içeren saksılara aktarıldı.
Dönüştürülmüş soya fasulyesi bitkilerinde GmDREB6 transgeninin varlığı PCR ile doğrulanmıştır. 10 transgenik soya fasulyesi bitkisinden alınan GmDREB6 transgeninin PCR ürünleri, GmDREB6 transgeninin boyutuna karşılık gelen yaklaşık 0.70 kb’lik bir bant gösterdi. T0 generasyonunda PCR testinde pozitif sonuçlar gösteren dokuz transgenik soya fasulyesi bitkisi T0-1, T0-2, T0-4, T0-5, T0-6, T0-7, T0-8, T0-9 ve T0-10. GmDREB6 transgeninin bu 9 GmDREB6-pozitif soya fasulyesi bitkilerinin genomuna dahil olup olmadığını doğrulamak için Southern blotlama gerçekleştirildi. Southern blotlama sonuçları, Şekil 2’de gösterilmektedir. Bu, DNA bantlarının dokuz GmDREB6-pozitif soya fasulyesi bitkisinin sekizinde gözlemlendiğini gösterirken, T0-6 ve transgenik olmayan (kontrol) bitkiler için böyle bir bant gözlenmediğini göstermektedir. T0-5, genin 2 kopyasına karşılık gelen 2 DNA bandı gösterirken, geri kalan çizgiler, T0-1, T0-2, T0-3, T0-4, T0-7, T0-8, T0-9 ve T0- 10, sadece 1 kopya gösterdi, bu da dönüşüm veriminin% 1.78 olduğunu gösteriyor.

Fotoğraf :T0 jenerasyonunun GM soya fasulyesi bitkilerinde GmDREB6 geninin varlığı, (A): geleneksel PCR ve (B): Southern blotting ile değerlendirilir. ↙: Kırmızı oklar GmDREB6 geninin varlığını gösterir. M: 1.0 kb DNA merdiveni; (+): pBI121 :: GmDREB6 vektörü; (-): WT: Yabani tip; 1–10: T0 jenerasyonunun GMsoya bitkileri T0-1, T0-2, T0-3, T0-4, T0-5, T0-6, T0-7, T0-8, T0-9 ve T0-10 olarak etiketlendi.
T0 jenerasyonunda başarılı bir şekilde dönüştürülen sekiz bitki arasında, T0-2, T0-4, T0-7, T0-9 ve T0-10 olmak üzere beş bitki, T1 neslinde tohumların yanı sıra çiçekler üretti ve T1-2, T1-4, T1-7, T1-9 ve T1-10 olarak etiketlendiler. T2 generasyonunda elde edilen tohumlar T2-2, T2-4, T2-7 ve T2-10 olarak etiketlendi.
Rekombinant DREB6 proteini, C terminalinde, endojen DREB6 proteininden farklı olan ve bir c-myc antikoru kullanılarak Western blotting yoluyla rekombinant DREB6 proteininin saptanmasına izin veren c-myc etiketine sahiptir. T1 generasyonunun transgenik soylarından alınan yapraklar, rekombinant DREB6 proteininin ekspresyonunu analiz etmek için kullanıldı. Western blotting analizinin sonuçları; 4 GM soya fasulyesi hattı olan T1-2, T1-4, T1-7 ve T1-10’da rekombinant DREB6 protein kDa’nın moleküler ağırlığına karşılık gelen yaklaşık 27’lik bir bant gösterdi; T1-9 çizgisinde ve transgenik olmayan soya fasulyesi bitkilerinde bant tespit edilmemiştir (Şekil 3). GM soya fasulyesi bitkilerinde rekombinant DREB6 protein ekspresyon analizinin sonuçları, GmDREB6 transgeninin bir nesilden diğerine aktarıldığını ortaya koydu.

Fotoğraf :T1 jenerasyonunun GM soya fasulyesi bitkilerinde rekombinant DREB6 proteini için Western blot. M: standart protein merdiveni; T1-2, T1-4, T1-7, T1-9 ve T1-10: T1 GM soya fasulyesi bitkileri; (-): WT; (+): C-myc etiketli> 100 kDa protein.
GM soya fasulyesi hatlarında ve transgenik olmayan bitkilerde GmP5CS geninin ekspresyonunun analizi GmP5CS gen ekspresyonunun analizi, ekspresyonun GM hatlarında hem normal hem de tedavi edilmiş koşullar altında transgenik olmayan bitkilerden daha yüksek olduğunu ortaya koymuştur (Şekil 4). Transgenik çizgilerdeki GmP5CS ekspresyon seviyesi, normal koşullar altında 1.36 ile 2.01 kat ve tuz stresi koşulları altında 1.58 ila 3.16 kat arasında değişmektedir. GmP5CS geninin ekspresyon seviyesi T2-7 çizgisinde en yüksek seviyedeydi (tuz stresi koşulları altında 3.16 kat). Duncan testi yapıldı ve sonuçlar, normal ve tuzla tedavi edilen koşullar altında GmDREB6 genini aşırı eksprese eden GM soya fasulyesi bitkilerinde ve transgenik soya hatlarında GmP5CS geninin ekspresyon seviyesindeki artışın önemli ölçüde (P <0.05) olduğunu ve transgenik olmayanda farklı şekilde eksprese edildiğini gösterdi.
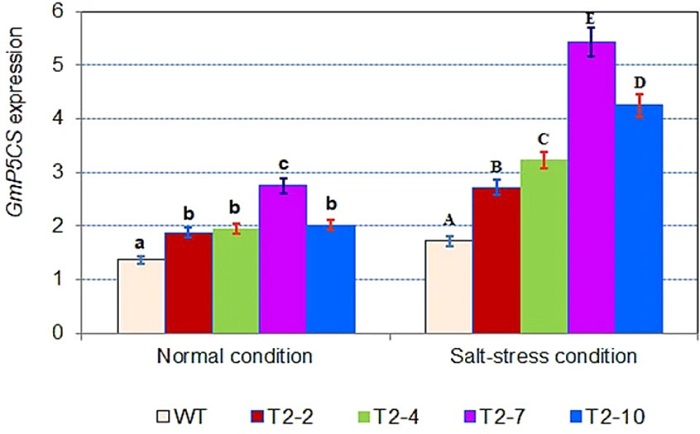
Fotoğraf : mP5CS geninin, T2 jenerasyonunun dört GM hattında ve qRT-PCR ile belirlenen transgenik olmayan bitkilerde (WT) ekspresyon paterni. Aktin (152 bp) referans gen olarak kullanıldı. WT: Yabani tip; T2-2, T2-4, T2-7 ve T2-10: T2 jenerasyonunda GM soya fasulyesi hatları. Sütunların üzerindeki farklı harfler, Duncan’ın testleriyle ölçüldüğü üzere istatistiksel olarak önemli farklılıkları (P <0.05) gösterir. Veriler, üç biyolojik kopya ± standart hatanın ortalamasını temsil eder.
GM ve transgenik olmayan soya fasulyesi bitkilerindeki prolin içeriğinin tuz stresi koşulları altında analizi dört GM soya fasulyesi hattının prolin içeriği, T2-2, T2-4, T2-7 ve T2-10 için bu bitkiler 7 gün boyunca tuz stresine tabi tutulduktan sonra analiz edildi. Sonuçlar prolin seviyelerinin GM bitkilerinde transgenik olmayan bitkilere göre daha yüksek olduğunu göstermiştir. Prolin içeriğindeki artış, normal koşullar altında büyüyen bitkilere kıyasla% 247.83 ila% 300.00 arasında değişmektedir. GM bitkileri arasında, T2-7 GM hattı tuz stresi koşulları altında en yüksek prolin içeriğine sahipti (P <0.001) (Tablo 1). Bu sonuçlar GM hatlarının transgenik olmayan bitkilerden daha yüksek bir tuz toleransına sahip olduğunu ve T2-7 hattının burada incelenen GM bitkileri arasında en yüksek tuz toleransına sahip olduğunu düşündürmekte.
| 7 günlük tuz stresine tabii tutulmuş GM fasülyelerinin Proline içeriği değişimi | |||||
| GM ve transgenik olmayan bitkiler | Normal Koşullardaki Proline içeriği (μmol/g)A | Proline içeriği (μmol/g)B | Normal durumla karşılaştırıldığındaki oran (%) | Transgenik olmayan bitkilerle karşılaştırıldığındaki oran (%) | |
| WT | 1.57a ± 0.027 | 3.74a ± 0.081 | 238.22 | 100.00 | |
| T2-2 | 1.84b ± 0.041 | 4.56b ± 0.099 | 247.83 | 121.93 | |
| T2-4 | 2.32c ± 0.034 | 6.16c ± 0.044 | 265.52 | 164.75 | |
| T2-7 | 2.59c ± 0.029 | 7.77d ± 0.069 | 300.00 | 207.75 | |
| T2-10 | 2.34c ± 0.056 | 6.29c ± 0.042 | 268.80 | 168.18 | |
(Tablo 1 : 2 gün boyunca tuz işlemine tabi tutulduktan sonra T2 jenerasyonunun GM soya fasulyesi hatlarının ve WT bitkilerinin prolin içeriği.)
Sesil organizma olarak, bitkilerin hayatta kalmak ve üremek için abiyotik çevresel koşulları algılamak ve bunlara yanıt vermek için bir sisteme sahip olması gerekir. Soya fasulyesi gibi önemli ekonomik ürünler için; başka bir görevden sorumlu bu abiyotik streslerden kaynaklanan strese karşı onları koruyup verimi artırır veya kayıpları azaltır. Genellikle, fizyolojik ve agronomik tepkiler, duyarlı genlerin ekspresyonunu tetikleyen moleküler anahtarlarla başlar. Bu genler arasında, DREB (dehidratasyona duyarlı eleman bağlayıcı proteinler), bitkinin başa çıkmasına yardımcı olan metabolik ve fizyolojik yanıtları tetikleyen, promotörlerin genleri üzerindeki belirli bölgelere bağlanan promotör genleri üzerindeki belirli bitki transkripsiyon faktörleridir (TF’ler). Daha önce kuraklık, ısı ve tuz gibi abiyotik streslerle, DREB’in, bitkilerin stres sinyal yollarında yer alan genlerin ekspresyonunu aktive etmek için bir A / GCCGAC motifi içeren DREB / CRT (C-tekrar) sekansına bağlanabilen bir trans-etkili faktör olduğu gösterilmiştir. DREB / CBF proteinlerinin tek bir 60 amino asit uzunluğunda DNA bağlayıcı AP2 alanı, kuraklık / soğuk / tuz stresine duyarlı promotör elemanlarını tek bir molekül olarak tanımasına ve bağlanmasına izin verir. Bu nedenle, DREB’ler / CBF’ler çok çeşitli bitkilerde tanımlanmış ve genetik olarak farklı promotörler kullanarak abiyotik gerilimlere daha yüksek toleranslı transgenik nesiller üretmek üzere tasarlanmıştır. Soğuk ve kuraklığa müdahale yollarında yüksek oranda yer alan DREB1 ve DREB2, birçok tesiste iyi bir biçimde tanımlanmıştır. Bununla birlikte, DREB6 genleri üzerindeki çalışmalar hala sınırlı. Bu çalışmada, soya fasulyesinde tuz toleransını artırmak amacıyla, soya fasulyesi genomunda bulunan DREB alt ailesinin bir üyesi olan GmDREB6 genini seçtik. Enéas ve arkadaşları, fasulye alt familyasındaki ortak DREB genini tanımladı ve ekspresyonlarını inceledi. Bu fasülye alf familyasından elde edilen ortak genomdan izole edilen 54 varsayılan PvDREB geni tanımlandı ve altı ana alt gruba ayrıldı. Ek olarak, dört PvDREB geni izole edildi ve dehidrasyon, yüksek tuzluluk oranı, düşük sıcaklık ve absisik asit tedavisine verdikleri cevaplar açısından analiz edildi. Bunlardan; kuraklık, tuz, soğuk ve ABA ile ilişkili PvDREB1F ve PvDREB5A’nın ifadesi; PvDREB2A ve PvDREB6B dehidrasyon ve soğuk koşullar altında indüklendi. Çalışma, fasulyede ortak kuraklık, tuz ve soğuk toleransı ile ilişkili moleküler mekanizmaların anlaşılmasına yardımcı olmuştur.
Abiyotik streslere bir diğer önemli cevap, bitki hücrelerindeki prolin içeriği ile yakından ilişkili olan ozmotik ayardır. Buna ek olarak; prolin bir ozmolit rolü oynamanın yanı sıra, abiyotik streslere, özellikle tuz ve kuraklık streslerine yanıt olarak metal bir selectör ve antioksidatif savunma molekülü olarak da hareket eder. Bitkilerin strese dayanıklılığı, tuz gibi stres koşulları ve su eksikliği koşulları altında düşük konsantrasyonlarda eksojen prolin uygulanarak güçlendirir. Prolin kodlayan P5CS geninin, abiyotik stres sırasında prolin birikimini indükleyen stres tepkisini düzenlemede kritik bir rol oynadığı bilinmektedir. Prolin birikimi, ozmotik basınçtaki artışın ana nedenlerinden biridir ve bu da bitkilerin su tutma yeteneğini artırır. Bitkilerdeki prolin biyosentez yolu; iki anahtar enzim olan P-pirolin-5-karboksilat redüktaz (P5CR) ve P-pirolin-5-karboksilat sentetazın (P5CS) katılımını içerir (Şekil 5). P5CS, GmP5CS geni tarafından kodlanır ve GmP5CS geninin promotör bölgesi, GmDREB6 TF’nin bağlanabildiği ve aktif gen ekspresyonunu yapabildiği GT-1 bölgesini içerir. Burada GM soya fasulyesi hatlarında ve özellikle T2-7 hattında tanımlandığı gibi P5CS geninin ekspresyonunda bir artışla indüklenen prolin birikimindeki bir artış, bitkileri oksidatif ve ozmotik streslere karşı koruyabilir. Önceki çalışmalar, P5CS’nin aşırı ekspresyonunun transgenik patates bitkileri, fasulye ve bitkisel soya fasulyesinde prolin birikimini arttırdığını göstermiştir. Hmida-Sayari ve arkadaşlarının araştırması, transgenik patates bitkilerindeki prolin içeriğinin kontrol bitkilerinden daha yüksek olduğunu göstermiştir. Dikkat çekici bir şekilde, transgenik patates bitkilerinde 100 mM NaCl’de prolin birikimi arttırıldı ve tuzluluk toleransı geliştirildi. Sonuç olarak, yumru verimi ve ağırlığı transgenik olmayan bitkilerden daha az azaltılmıştır. Chen ve arkadaşları, Vigna aconitifolia‘daki P5CS ekspresyonu üzerinde çalışmış ve tuz stresi tedavisinde 48 saat sonra kontrol bitkilerine kıyasla prolin içeriği içeren genetik olarak modifiye edilmiş bitkiler üretmişlerdir. Zang ve diğerleri (2015) soya fasulyesinde aşırı P5CS genini eksprese etmiş ve yabanıl tip bitkilerden daha yüksek tuz toleransı ve daha yüksek prolin içeriğine sahip T2, T3 transgenik soyları elde etmiştir.
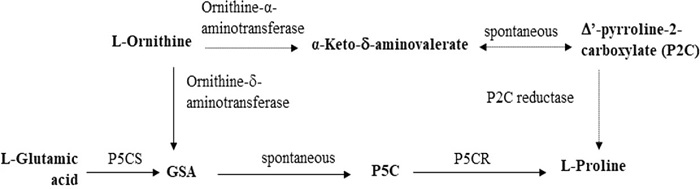
Fotoğraf : Bitkilerde prolin biyosentez yolları.
Kuraklık, soğuk ve tuz stresleri altında fasulye içindeki PvP5CS’nin ekspresyon paternleri üzerinde yapılan araştırmalar, bu abiyotik streslerin fasulye yapraklarında PvP5CS ekspresyonunu belirgin şekilde arttırdığını gösterdi. Tuz işleminden sonra fasulye bitkilerindeki PvP5CS’nin transkripsiyonel ekspresyon seviyelerinin sonuçları, tedaviden 2 saat sonra 16.3 kata kadar daha yükseldi. Ek olarak, prolin birikimi 2 saatte 1.3 kattan, 6 saatlik tedavilerde 2.7 kat yükselmiştir.
Bu çalışmanın sonuçları, GmDREB6 geninin aşırı ekspresyonunun, tuz stresi koşulları altında büyüyen bitkilerde daha yüksek hücre içi prolin içeriğinden sorumlu olan GmP5CS geninin transkripsiyon seviyelerini arttırdığını göstermiştir (Tablo 1).
Bu nedenle soya fasulyesinde GmDREB6 geninin aşırı ekspresyonunun; stres tepkisinde önemli bir rol oynayan prolin birikimiyle sonuçlanan P5CS geninin transkripsiyonel ekspresyonunu arttırarak tuz toleranslarını arttırdığını düşünüyoruz.
35S-GmDREB6-c-myc yapısı, Agrobacterium aracılı transformasyon ile soya fasulyesine başarıyla sokuldu ve T2 jenerasyonuna kadar GM hatları üretildi. GmDREB6 geninin ekspresyon seviyeleri ve prolin birikimi T2 jenerasyonunda arttı ve normal ve tuzla işleme tabi tutulan koşullar altında vahşi tip (WT) bitkilerinden daha yüksekti. Bu çalışmanın sonuçları, GM soya fasulyesi bitkilerinde prolin birikiminin arttırılmasında GmDREB6 geninin aşırı ifadesinin etkinliğini gösterdi ve GmDREB6’nın, çapraz konuşma tepkisi nedeniyle tuz ve kuraklık streslerine karşı toleransı artırmak için genetik mühendisliğinde potansiyel bir aday olarak hareket edebileceğini gösterdi.
DT84 soya fasulyesi çeşitlerinden tohumlar Vietnamdan Vietnam Tarımsal Genetik Enstitüsü tarafından sağlandı. PBI121 :: GmDREB6 ifade vektörünü taşıyan rekombinant A. tumefaciens CV58 suşu, Thai Nguyen Eğitim Üniversitesi, Vietnam Biyoloji Okulu, Genetik ve Modern Biyoloji Bölümü tarafından sağlandı. GmDREB6-c-myc yapısı, 693 bp kodlama bölgesi, XbaI için kesme bölgesini içeren 5 uçta 8 bp segment (GCTCTAGA) ve 3’te 7 bp segment (GAGCTCG) içeren GmDREB6 genini içerir. I SacI için kesme bölgesini içeren uç. C-myc antijenini kodlayan 33 bp’lik bir segment, 3-ucuna eklenmiştir (Şekil 6). Bu, Agrobacterium aracılı transformasyon ile soya fasulyesi kotiledonunu dönüştürmek için kullanıldı. Bu vektör, kurucu promotör CaMV 35 S’nin (karnabahar mozaik virüsü) kontrolü altındadır. Kaset yapısında iki işaretleyici gen de vardı: antibiyotik kanamisine direnç veren ve transgeni içeren kolonileri seçmek için kullanılan nptII geni (neomisin fosfotransferaz II); ve Batı lekesindeki hedef proteinin saptanması için bir antijen olarak işlev gören c-myc geni (c-myc peptidini kodlayan) kullanılmıştır.
| Primer | Nükleotit sekansı(5’-3’) | Boyut |
| GmREB6-F/GmDREB6-R | CATAGAAGAAGCCACTAACACTACA | 741bp |
| ATTCAGATCCTCTTCTGAGATGAGT | ||
| nptII-F/nptII-R | GAGGCTATTCGGCTATGACTG | 983 bp |
| ATCGGGAGCGGCGATACCGTA | ||
| qGmP5CS-F/qGmP5CS-R | CGAACTGAGCTTGCAGAGGGGC | 165 bp |
| TCGCTTAGCCTCCTTGCCTCC | ||
| qAct-F/qAct-R | CCTAGCATTGTTGGTCGTCCTC | 152 bp |
| CATATCATCCCAGTTGCTAACAAT |
(Tablo 2: PCR’de ve DNA problarının sentezi için kullanılan primer çiftlerinin nükleotit sekansları.)
Agrobacterium aracılı transformasyon için kullanılan pBI121 :: GmDREB6 vektörünün diyagramatik gösterimi. LB: sol T-DNA sınırı; RB: sağ T-DNA sınırı; nptII: neomisin-fosfo-transferaz II; CaMV35S: karnabahar mozaik virüsü 35S promoteri; Soya fasulyesi bitkilerinin mRNA’sından izole edilen GmDREB6 geni; c-myc: c-myc peptidini kodlayan nükleotid dizisi. Nospro: nopalin sentaz promotörü; Noster: nopalin sentaz terminatörü; Hind III, Xba I ve Sac I için kısıtlama bölgeleri düz siyah çizgilerle gösterilir.
Soya fasulyesinin kotiledon düğümünden agrobacterium aracılı transformasyon ve soya fasulyesi bitkilerinin rejenerasyonu, daha önce Olhoft ve arkadaşları tarafından tarif edilen metodolojiye göre gerçekleştirildi. Sterilize edilmiş soya fasulyesi tohumları Murashige ve Skoog (MS) ortamında çimlenmiştir. 4 gün sonra dönüştürülecek materyal olarak kotiledonlar kullanıldı. Kotiledonlar 30 dakika boyunca bakteriyel süspansiyona daldırılmış ve daha sonra antibiyotik olmayan bir ortak kültür ortamına (CCM) aktarılmıştır. Dönüştürülen numuneler, 400 mg / L sefotaksim içeren bir sürgün indüksiyon ortamında (SIM) 10 dakika süreyle yıkandı ve SIM’de 50 mg / L kanamisin (1. kez) ile kültürlendi. 2 hafta sonra örnek, 75 mg / L kanamisinde (2. kez) SIM’e aktarıldı. Seçilen ortam üzerinde hayatta kalan sürgün kümeleri kotiledonlardan çıkarıldı ve 50 mg / L kanamisin içeren sürgün büyütme ortamına (SEM) aktarıldı. Sürgünler yaklaşık 3-4 cm uzunluğa kadar büyüdüğünde, tam bir bitkinin oluşumu için 50 mg / L kanamisin içeren bir köklendirme ortamına (RM) transfer edildi. Sağlıklı bitkiler 1: 1 oranında “pirinç kabuğu külü: kum”içeren saksılara aktarıldı. Yaklaşık 1-2 hafta sonra, hayatta kalan bitkiler bir seraya transfer edildi. İn vitro koşullar altında transforme kotiledonlardan rejenere edilen soya fasulyesi bitkileri T0 generasyonu olarak adlandırıldı. T0 generasyonunun bitkilerinden elde edilen tohumlar T1 generasyonu olarak kabul edildi ve T1 bitkilerinden elde edilen tohumlar T2 generasyonunu temsil etti.
Toplam DNA, Saghai-Maroof ve arkadaşları tarafından tarif edilen yönteme göre rejenere soya fasulyesi bitkilerinin genç yapraklarından izole edildi ve daha sonra% 0.8 agaroz jel üzerinde incelendi. Rejenere soya fasulyesi bitkilerinde GmDREB6 transgen varlığını doğrulamak için PCR yapıldı. PCR primerleri SoyDREB6F / SoyDREB6R’nin nükleotit sekansları Tablo 2’de listelenmiştir. PCR, 20 uL’lik son olarak belli bir hacim kullanılarak gerçekleştirildi ve her primerden 0.5 μL (10 pmol / µL), 12.5 mL 2x Ana karışım, 2.0 uL cDNA (500 ng / mL) ve 4.5 uL su içeriyordu. Termosiklin koşulları ile ayarlanan PCR, aşağıdaki aşamaları içermektedir: 5 dakika boyunca 95 ° C’de bir ilk denatürasyon, ardından 20 saniye boyunca 95 ° C’de, 20 saniye boyunca 58 ° C’de 35̊C çevrim ve 30 saniye boyunca 72 ° C ve 10 dakika boyunca 72 ° C’de aşamaları ile ısı silkulasyonuna maruz bırakıldı. PCR ürünleri,% 1.0 agaroz jelinden eksize edildi ve talimatlara göre bir jel ekstraksiyon kiti (Qiagen, Venlo, Hollanda) ile saflaştırıldı.
GmDREB6 transgenini ihtiva eden ve bu nedenle PCR ile başarılı bir şekilde amplifiye edilen GM bitkilerinin DNA’sı, Southern blot analizine tabi tutuldu. Analiz, pBI121 :: GmDREB6 vektörü içinde nptII geninin varlığını saptamak için yapıldı. PCR ile başarılı bir şekilde amplifiye edilen T0 generasyonunun GM soya fasulyesi bitkilerinden genomik DNA örnekleri, 37 ° C’de SacI kısıtlama enzimi kullanılarak gece boyunca bekletildi. Bundan sonra, numuneler% 1.0 agaroz jeli üzerinden alınarak bir selüloz membrana aktarıldı. PBI121 :: GmDREB6 vektörü içindeki nptII geni, nptII-F / nptII-R PCR primerleri kullanılarak amplifiye edildi (Tablo 2) ve prob, bir Biotin DecaLabel DNA Etiketleme Kiti kullanılarak biyotin-11-dUTP ile etiketlendi.
Toplam protein, T1 jenerasyonunun pozitif olarak transforme edilmiş GM soya fasulyesi bitkilerinin yapraklarından ekstrakte edildi ve Laemmli25 tarafından tarif edildiği gibi% 10 jel kullanılarak SDS-PAGE’e tabi tutuldu. Daha sonra numuneler Western blot analizi için nitroselüloz membranlara aktarıldı. Membranlar daha sonra gece boyunca bloke edici çözelti (Fosfat Tamponlu Salin (PBS) – Tween içinde% 5 yağsız süt) içerisinde bloke edildi ve birincil antikor (c-myc) ile inkübe edildi. Daha sonra numuneler 3 saat çalkalandı, ardından PBS (1x) ile üç kez yıkandı ve 2 saat boyunca ikincil bir antikor ile inkübe edildi. C-myc ile konjüge edilmiş fare monoklonal antikoru, 1: 700’de PBS içinde% 5 süt ile seyreltilerek birincil antikor olarak kullanılırken, Bayır turpunun peroksidazı ile konjüge edilmiş 1: 4000’de PBS içinde% 5 süt ile seyreltildi ve ikincil antikor olarak kullanıldı. Sonuçlar 3,3’ ,5,5’ tetrametilbenzidin ile gözlendi.
T2 generasyonunun tohumları, yani T2-2, T2-4, T2-7 ve T2-10 ve WT bitkilerinin tohumları, 23 ° C’lik sıcaklıkta, nokturnalde gölgeli bir serada çimlendirildi ve büyütüldü. (16 saat / 8 saat ışık / karanlık sirkulasyonu altında 20 ° C sıcaklıkta.) Üç yapraklı aşamada (V3), T2 generasyonunun GM bitkilerine yedi gün boyunca 0 (kontrol), 150 ve 300 mM’lik farklı konsantrasyonlarda sodyum klorür (NaCl) çözeltileri ile muamele edildi. Bitkiler her 2 günde bir 3 farklı NaCl konsantrasyonu ile sulandı. Bu nedenle, kontrol düzeneğindeki bitkiler NaCl (0 mM) içermeyen düz su alırken, deneysel bitkilerin tümü üç kez NaCl çözeltisi ile sulandı; ilk kez 150 mM’lik bir konsantrasyonda NaCl çözeltisi ile 1. günde, ikinci ve üçüncü kez 300 mM’lik bir konsantrasyonda NaCl çözeltisi ile, sırasıyla 3. ve 5. günde muamele gördü. Deney ve kontrol bitkilerinin yaprakları, deneyin başlangıcından itibaren 7. günde toplandı, ardından P5CS geninin ve prolin içeriğinin transkripsiyonel seviyesini belirlemek için analizde kullanıldı.
GM ve WT soya fasulyesi bitkilerindeki GmP5CS geninin, normal ve tuz ile işleme tabi tutulmuş koşullar altında ekspresyon seviyesi, SYBR Green I floresan boya kullanılarak kantitatif olarak analiz edildi. T2 jenerasyonunun yapraklarından elde edilen toplam RNA, TRIzol kiti ile ekstre edildi ve cDNA, üreticinin talimatlarına göre First Strand cDNA Sentez Kiti kullanılarak sentezlendi. Aktin geni (SAc1 geni, GenBank erişim numarası: J01298.1) normalizasyon için referans olarak kullanıldı. QRT-PCR testi, her primerden 0.5 μL (10 pmol / µL), 12.5 mL 2x Ana karışım, 2.0 uL cDNA (500 ng / mL) ve 4.5 uL su içeren 20 μL’lik bir hacim kullanılarak gerçekleştirildi. Termosiklin koşulları ile belirlenen qRT-PCR, aşağıdaki aşamaları içermektedir: 10 dakika boyunca 95 ° C’de bir ilk denatürasyon, ardından 10 saniye boyunca 95 ° C’de, 10 saniye boyunca 58 ° C’de 45 ° C sıcaklık döngüsü ve 20 saniye boyunca 72 ° C’de, şeklinde ısı sirkulasyonundan sonra analiz edildi. Akış sıcaklığı 1 dakika boyunca 65-95 ° C’den arttığında, floresan sinyalleri toplandı. Gen ekspresyon seviyeleri Livak ve Schmittgen tarafından tarif edilen R = 2 −Ct yöntemi kullanılarak nicelendi. Sonuçlar aktin kodlayan genlerin sonuçlarına göre sunulmuştur.
Daha sonra normal ve tuz ile muamele edilmiş koşullar altında büyüyen GM ve WT soya fasulyesi bitkilerinin prolin içeriği analiz edildi. GM bitkilerinin prolin içeriği Bates ve arkadaşları tarafından tarif edilen yöntemle belirlenmiştir. Transgenik ve transgenik olmayan bitkilerin yapraklarındaki prolin seviyeleri, yedi gün boyunca tuz stresine maruz bırakıldıktan sonra analiz edildi. Yaklaşık 0.5 g bitki materyali, 10 ml% sulandırılmış sülfosalisilik asit içinde homojenleştirildi ve homojenat, 12000 rpm’de 10 dakika süreyle santrifüjlendi. 2 mL süzüntü içeren bir karışım, 100 ° C’de 1 saat boyunca bir deney tüpünde 2 mL asit ninhidrin ve 2 mL buzlu asetik asit ile reaksiyona sokuldu. Reaksiyon bir buz banyosunda tutuldu ve kromofor, 4 mL tolüen ile özümlendi; bunu takiben absorbansı, bir BioMate spektrofotometre kullanılarak 520 nm’de belirlendi. Prolin içeriği taze kütlenin μmoL / g olarak ifade edildi. Elde edilen tüm veriler SPSS yazılımı kullanılarak tek yönlü varyans analizine tabi tutuldu. Veriler Duncan’ın testi ile analiz edildi (P = 0.05, 0.001).
Kaynak : nature.com



